 Elizabeth Martin-Silverstone* describes her investigations into pterosaur flight, and the evolution of flying vertebrates
Elizabeth Martin-Silverstone* describes her investigations into pterosaur flight, and the evolution of flying vertebrates
Thanks to the Society’s Daniel Pigeon Fund, I recently spent a few weeks travelling around Germany in search of pterosaur fossils in various collections.
Picture: An azhdarchid pterosaur model greets visitors to the Staatliches Museum für Naturkunde Karlsruhe (SMNK).
Flight is a truly remarkable adaptation that has evolved only three times in vertebrates: in birds, bats, and in the extinct flying reptiles that are the subject of my studies - the pterosaurs. Of all these different lineages, pterosaurs produced the widest range of body sizes (with wing-spans ranging from 0.2 to 11m!), the largest overall size (the largest extant bird has a wingspan of 4 m, largest extinct was 7m) yet were also the first to achieve powered flight, first in the Late Triassic - picture below. Despite being known from the fossil record for over 200 years, basic questions on pterosaur biology and biomechanics - such as reliable body mass estimates and flight abilities - remain unknown.
Body mass
Estimating body mass is one of the most basic and essential features in understanding locomotory capabilities of an animal, and is even more important with respect to flying animals, because an animal’s mass is directly proportional to its ability to take off and achieve lift. As the largest animals ever to take to the air, pterosaurs pushed the limits of aerodynamics, and as there remains much debate around whether or not these large pterosaurs could actually fly, making accurate mass estimates vital to our overall comprehension of them. Previous mass estimates have varied drastically depending on the methods used. For example, one of the largest pterosaurs (Quetzalcoatlus northropi) has been estimated at an improbably light 70kg1, at a more reasonable 250kg2 and very heavy 544kg3.
Fortunately, modern imaging technology allows us to make accurate estimates of bone mass in relatively uncrushed specimens using computed tomography (CT) scans4. These take x-rays from several different angles, allowing the fossil to be visualised in 3D, and the volume of actual bony material within each element to be calculated. When multiplied by bone density, this gives you a mass estimate.
 Picture: Artist’s impression of an azhdarchid pterosaur Arambourgiania showing just how big they were when compared with a giraffe and a human. Image by Mark Witton, used with permission.
Picture: Artist’s impression of an azhdarchid pterosaur Arambourgiania showing just how big they were when compared with a giraffe and a human. Image by Mark Witton, used with permission.
Finally, after the bones have been digitally visualised, muscle mass can be estimated by reconstructing the muscle attachments and muscle volumes. This should produce a more accurate estimate of total body mass. This is what I am working towards now. CT scans also give us an idea of the degree of ‘pneumaticity’, which allows for comparison between different elements and species through the bone’s Air Space Proportion (ASP). This can also be performed using cross-sections, but these are less accurate5].
Pneumaticity, which is the presence of air sacs and diverticulae that invade the soft tissue and bones as part of the animal’s respiratory system, is also found in birds, some theropod dinosaurs, and sauropod dinosaur necks. ASP gives a quantifiable measurement of the pneumaticity, which has implications for the biomechanics of an animal as it directly affects the stiffness of a bone.
Zittel wing
So far, I have visited the collections of four German museums: Staatliches Museum für Naturkunde Karlsruhe (SMNK), Staatliches Museum für Naturkunde Stuttgart (SMNS), Institut für Geologie und Paläontologie, Universität Tübingen (GPIT), and the Bayerische Staatssammlung für Paläontologie und Geologie, Munich (BSPG). I have been able to take a large number of pictures to start understand the variation in ASP throughout the skeleton and in different species
 I greatly enjoyed the museum visits - especially Karlsruhe, where I was greeted at the front of the museum by a giant azhdarchid model and got to stay there (which felt a bit like Night at the Museum). There’s also a giant fuzzy azhdarchid model in flight hanging from the ceiling of the museum! (picture, left). Munich was fascinating for the number of historically significant specimens in its collection, including the first pterosaur known to science (unfortunately that one is highly protected and I wasn’t able to work on it). I was completely surprised when I opened a drawer to discover the ‘Zittel wing’, the famous fossil that first gave us an idea of pterosaur wing structures, way back in 1880.
I greatly enjoyed the museum visits - especially Karlsruhe, where I was greeted at the front of the museum by a giant azhdarchid model and got to stay there (which felt a bit like Night at the Museum). There’s also a giant fuzzy azhdarchid model in flight hanging from the ceiling of the museum! (picture, left). Munich was fascinating for the number of historically significant specimens in its collection, including the first pterosaur known to science (unfortunately that one is highly protected and I wasn’t able to work on it). I was completely surprised when I opened a drawer to discover the ‘Zittel wing’, the famous fossil that first gave us an idea of pterosaur wing structures, way back in 1880.
Picture, right: The famous ‘Zittel Wing’ Rhamphorhynchus specimen from the BSPG in Munich, known for the well preserved complete wing bones and soft tissue showing the shape of the membrane wing.
Trends
 While the work is still very much in its infancy, some trends are becoming visible. First of all, ASP is consistently lower (meaning that there is proportionally more bone than air), in smaller pterosaurs (e.g. Rhamphorhynchus, Pterodactylus, Germanodactylus) than larger ones (e.g. Ornithocheirus, Anhanguera, azhdarchid pterosaurs). Previous studies have suggested that pterosaurs are among the most, if not the most, pneumatic animals to have lived5. However, this now appears to be true of only the larger pterosaurs, and is not the case for smaller ones.
While the work is still very much in its infancy, some trends are becoming visible. First of all, ASP is consistently lower (meaning that there is proportionally more bone than air), in smaller pterosaurs (e.g. Rhamphorhynchus, Pterodactylus, Germanodactylus) than larger ones (e.g. Ornithocheirus, Anhanguera, azhdarchid pterosaurs). Previous studies have suggested that pterosaurs are among the most, if not the most, pneumatic animals to have lived5. However, this now appears to be true of only the larger pterosaurs, and is not the case for smaller ones.
As ASP refers to the physical amount of air present within a bone, (the degree of hollowness if you will) this can tell us something about biomechanics. Pterosaur bones probably underwent deformation in flight (as all bones do in movement), and would have been particularly susceptible to bending. The bending resistance of a hollow tube, related to the second moment of area, is dependent on the amount of material in the tube and how far from the centre it is (how large of a diameter the bone has). Pterosaur bones commonly have cortical thicknesses of 0.5-2mm. For two bones with a cortical thickness of one millimetre, the bending stiffness is much higher (i.e., it is more resistant to bending) in a bone with a diameter of 30mm than one with a diameter of 10mm.
 Most pterosaur wing bones studied so far have ASP values ranging from 0.4-0.8, with very few exceeding 0.8. However, one pterosaur is showing much lower values, of just 0.1. This value was obtained from the dsungaripterid pterosaur “Lonchognathosaurus” from China. Dsungaripterids have been noted by pterosaur palaeontologists as having unusually thick-walled bones, which also affects their ASP. Modern birds that spend a significant time in water (such as cormorants, or penguins) have similarly thick-walled features, and dsungariperid palaeoecology is not fully understood. The implications of this thick-walled, low ASP feature are currently under investigation.
Most pterosaur wing bones studied so far have ASP values ranging from 0.4-0.8, with very few exceeding 0.8. However, one pterosaur is showing much lower values, of just 0.1. This value was obtained from the dsungaripterid pterosaur “Lonchognathosaurus” from China. Dsungaripterids have been noted by pterosaur palaeontologists as having unusually thick-walled bones, which also affects their ASP. Modern birds that spend a significant time in water (such as cormorants, or penguins) have similarly thick-walled features, and dsungariperid palaeoecology is not fully understood. The implications of this thick-walled, low ASP feature are currently under investigation.
Picture: Cross-sectional image of a smaller pterosaur wing phalanx (Rhamphorhynchus, SMNS 59421). Scale markings indicate 1cm.
The main purpose of this study was to CT scan as many pterosaur fossils as possible to understand pterosaur bone and body mass. This is going ahead thanks to the large number of specimens on loan from the SMNK, including one nearly complete Tapejara 6, two partial ornithocheirid wings, one partial Coloborhynchus skeleton, and more. So far, only portions of the Tapejara have been scanned, but these have been very successful (below right).
Future work
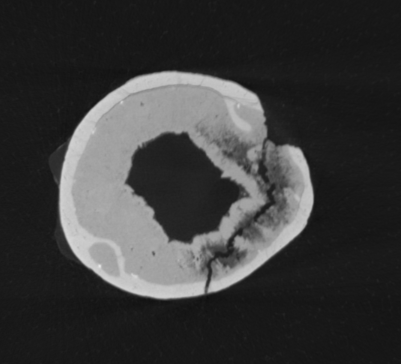 Picture: CT scan showing the cross-section of a long bone of Tapejara wellnhoferi (SMNK PAL 1137). Cortical bone is seen around the outside as white, while trabecular bone is visible as thinner white bone coming off of the thicker outer cortex (bottom left).
Picture: CT scan showing the cross-section of a long bone of Tapejara wellnhoferi (SMNK PAL 1137). Cortical bone is seen around the outside as white, while trabecular bone is visible as thinner white bone coming off of the thicker outer cortex (bottom left).
The main thing I have focused on during the last six months has been finishing the CT-scanning of all the SMNK specimens, and begin analysis - as the fossils need to be returned to Germany this month. I will also continue looking at the ASP, and how that affects the biomechanics of the pterosaur wing.
Depending on how much it will cost to return the specimens, I may visit additional museums in Germany as well, to collect yet more data.
Acknowledgements
In addition to the Society’s Daniel Pigeon Fund, additional funding for my PhD is provided by the Graduate School of the National Oceanography Centre, Southampton, and the Natural Sciences and Engineering Research Council of Canada (NSERC). I would like to thank all those who helped me at each museum and allowed me into their collections - Dino Frey (SMNK), Philipe Havlik (GPIT), Oliver Rauhut (BSPG) and Rainer Schoch (SMNS), and my supervisors for pushing me and helping me get to Germany – Gareth Dyke, Colin Palmer, Mike Habib, and Emily Rayfield.
- If you would like to find out more about Society fieldwork research grants, visit Research Grants. Applications for 2016 will be invited in September.
* University of Southampton
References
- Chatterjee S, Templin R (2004) Posture, locomotion, and paleoecology of pterosaurs: Geological Society of America Special Publications 376: 1-64.
- Witton MP (2008) A new approach to determining pterosaur body mass and its implications for pterosaur flight. Zitteliana B28: 143-159.
- Henderson DM (2010) Pterosaur body mass estimates from three-dimensional mathematical slicing. Journal of Vertebrate Paleontology 30: 768-785.
- Martin EG, Palmer C (2014) A novel method of estimating pterosaur skeletal mass using computed tomography scans. Journal of Vertebrate Paleontology 34: 1466-1469.
- Martin EG, Palmer C (2014) Air Space Proportion in Pterosaur Limb Bones Using Computed Tomography and Its Implications for Previous Estimates of Pneumaticity. PloS one 9: e97159.
- Eck K, Elgin RA, Frey E (2011) On the osteology of Tapejara wellnhoferi Kellner 1989 and the first occurrence of a multiple specimen assemblage from the Santana Formation, Araripe Basin, NE-Brazil. Swiss Journal of Palaeontology 130: 277-296.