Burrowing animals helped kill off ancient seafloor matgrounds. Catherine Mascord reports on the role of shallow burrowers with links to the oxygenated surface in initiating this process
Mascord C., The fossils of Bell Island.
Geoscientist 29 (2), 12-17, 2019
https://doi.org/10.1144/geosci2019-009;
Download the pdf here
On the surface, Bell Island, Newfoundland appears unremarkable in geological terms. The island is relatively flat with few hills or other dramatic geography, and most of the rocky outcrops are limited to the cliffs along the island’s coastline.
Throughout much of the early 20th century the island was host to one of the most productive iron ore mines in Canada. Today the mines are decommissioned, but the island’s rocks still hold a great deal of scientific interest because they preserve a record of one of the most important events in the history of life on Earth, the Cambrian Substrate Revolution. The revolution saw the transition from a seafloor lined with microbial mats and limited burrowing by animals, to more extensive vertical burrowing, oxygenation of the subsurface and the evolution of the animal-dominated seafloor we know today.
To better understand this record, I spent two weeks of fieldwork, in June 2018, examining the island’s geology, using the research funding provided by the Geological Society. From the information gathered during the trip it seems that shallow burrowing animals were key to opening up the ocean substrate to more complex animals.
 Left, Wet weather typified most of the fieldwork, however we had a few sunny days that showed off the crystal-clear waters and the glorious sedimentology in the cliffs themselves. The cliffs were not always accessible without the use of a boat or a lot of scrambling down near-vertical scree slopes. This particular beach, Freshwater Cove, was only accessible by climbing down a stream bed and using an abandoned fence (visible to the left) someone had carefully propped up against the cliff as a makeshift ladder.
Left, Wet weather typified most of the fieldwork, however we had a few sunny days that showed off the crystal-clear waters and the glorious sedimentology in the cliffs themselves. The cliffs were not always accessible without the use of a boat or a lot of scrambling down near-vertical scree slopes. This particular beach, Freshwater Cove, was only accessible by climbing down a stream bed and using an abandoned fence (visible to the left) someone had carefully propped up against the cliff as a makeshift ladder.
Subsurface reform
For much of Earth’s history life on the marine seafloor was very different to the animal-dominated communities we know today.
A typical healthy, modern seafloor is home to a large number of burrowing organisms. These animals, particularly those capable of producing deep burrows, are vital for maintaining oxygen levels in the sediment and recycling deposited material, because they mix oxygen and suspended material from the water column into the sediment as they burrow (McIlroy & Logan,
PALAIOS 1999). However, most burrowing animals did not evolve until the early to mid-Cambrian, about 530 million years ago. The few burrowers around in the Precambrian were rarely capable of burrowing more than a few millimetres into the surface sediment (Gehling, PALAIOS 1999; Droser
et al., PNAS 2002; Pecoits
et al., Science 2012).
Without the oxygenating effects of burrowing animals, the ancient seafloor was characterised by widespread anoxia (a low oxygen concentration) (McIlroy & Logan,
PALAIOS 1999) and in the place of animals, the seafloor was blanketed in a community of single-celled organisms known as a microbial matground. Matgrounds are formed when bacteria and other microbes stick themselves, and the host sediment together, using a biological glue (Parsons
et al., Geophys. Res. Lett. 2016), allowing them to form a cohesive mass on the sediment surface. Despite the microscopic size of their constituents, matgrounds, if left undisturbed, can reach thicknesses of a few cm or more. By gluing themselves together, the bacteria also increase their resistance to erosion and can further facilitate sediment anoxia by limiting sediment mixing by wave action.
Matground traces or microbial-induced sedimentary structures, such as stromatolites, elephant skin fabrics and wrinkle structures, are relatively common in the Precambrian fossil record. However, between 540 and 530 million years ago there was rapid transition from a matground-dominated seafloor to oxygenated, animal-dominated conditions. This transition—the Cambrian Substrate Revolution—is preserved in the fossil record as a rapid decline in fossilised microbial fabrics, and an increase in the number and diversity of trace fossils (Fedonkin,
Paleont. Jour. 1978; Crimes & Droser,
Ann. Rev. Ecol., Evol. & Syst. 1992; Jensen,
Int. & Comp. Biology 2003).
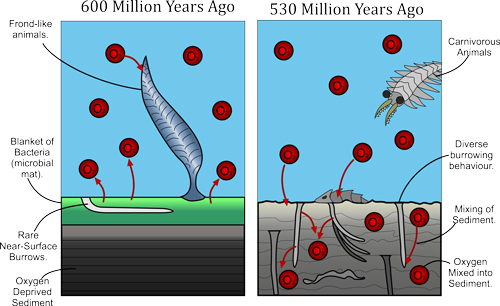 Right, The change in the seafloor environment from the Precambrian to the Cambrian. In the Precambrian (late Ediacaran 580-540 million years ago) the seafloor was blanketed by a microbial mat, with the few burrowing animals present only able to burrow into the top few millimetres of sediment. By the middle Cambrian (about 510 million years ago), the matgrounds had been replaced by a diverse community of burrowing organisms that mixed oxygen and other dissolved material into the sediment. This diverse animal-dominated community is characteristic of the seafloor today
Right, The change in the seafloor environment from the Precambrian to the Cambrian. In the Precambrian (late Ediacaran 580-540 million years ago) the seafloor was blanketed by a microbial mat, with the few burrowing animals present only able to burrow into the top few millimetres of sediment. By the middle Cambrian (about 510 million years ago), the matgrounds had been replaced by a diverse community of burrowing organisms that mixed oxygen and other dissolved material into the sediment. This diverse animal-dominated community is characteristic of the seafloor today
 Left, The fossils and trace fossils preserved in the Precambrian and Cambrian are very different from one another. Precambrian (Ediacaran) fossils (left) include simple Rangeomorph (frond-like) animals, shallow surface trails and elephant skin textures produced by microbial mats on the sediment surface. By the Cambrian (right) the Rangeomorphs had disappeared, replaced by other, more complex animals such as Trilobites. The trace fossils were also more complex because animals capable of deep burrowing had evolved
Left, The fossils and trace fossils preserved in the Precambrian and Cambrian are very different from one another. Precambrian (Ediacaran) fossils (left) include simple Rangeomorph (frond-like) animals, shallow surface trails and elephant skin textures produced by microbial mats on the sediment surface. By the Cambrian (right) the Rangeomorphs had disappeared, replaced by other, more complex animals such as Trilobites. The trace fossils were also more complex because animals capable of deep burrowing had evolved
From the late Cambrian and to the present-day, matground communities are rare, limited to anoxic, nutrient-deficient or polluted areas of the sea and lakes, where burrowing animals can’t survive in large numbers. This is partly because burrowing animals will break a matground apart as they burrow, and can feed on the microbes themselves, killing off the microbial community. However, matground-dominated seafloors, and thus the microbial-induced fabrics they create, do see a few resurgences in the fossil record after mass extinctions (Gerbersdorf
et al., J. Soils & Seds. 2009; Mata & Bottjer,
Geobiology 2012), most notably after the End-Permian mass-extinction (Feng
et al., Earth Sci. Rev. 2018; Buatois
et al., Geobiology 2013), where oceanic oxygen stagnation (Hotinski
et al., Geology 2001) likely led to a die-off of burrowing animals allowing matgrounds to thrive.
Understanding the mechanisms behind matground resurgence and decline could prove important to future conservation efforts, as global climate change has led to an increase in oceanic dead-zones that may allow matgrounds to regain a foothold today. However, matground resurgence and decline, particularly in the context of the Cambrian Substrate Revolution, is poorly understood. For example, it is unclear how early burrowing animals, which are likely to have lacked many of the more complex adaptations to anoxia, survived in the anoxic sediments of the Precambrian.
Extraordinary interaction
Most of Bell Island is made up of interlayered sandstones, mudstones and thick ironstones, deposited in a shallow sea just after the Cambrian Substrate Revolution, 490 to 470 million years ago, during the Upper Cambrian and into the Early Ordovician. Like many sedimentary rocks deposited in marine environments they preserve ripples and cross bedding produced by wave and current action.
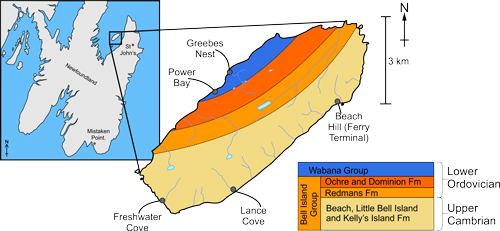 Right, Located in Northern Newfoundland, Bell Island is part of the Wabana and Bell Island Groups, dating from 490 to 470 million years ago. The geology is based on the published map by King, A.F. (1988).
Right, Located in Northern Newfoundland, Bell Island is part of the Wabana and Bell Island Groups, dating from 490 to 470 million years ago. The geology is based on the published map by King, A.F. (1988).
What makes Bell Island important to the understanding of the Cambrian Substrate Revolution is the fossil community it preserves. These fossils are not the shells and bodily remains you would normally think of when fossils are mentioned—indeed, Bell Island has surprisingly few body fossils—instead, the island’s rocks are characterised by a diverse community of trace fossils (fossils produced by the behaviour of an organism).
Bell Island’s trace fossil communities are unusual because they preserve both complex fossil burrows and microbial-induced sedimentary fabrics in the same succession and on the same bedding planes, recording an ancient community where both complex animals and matgrounds lived together. This rock archive therefore presents a unique opportunity to examine the Cambrian Substrate Revolution because it allows us to study an in-situ interaction between a well-established Precambrian-like matground and relatively complex burrowing animals—something that is rare in nature today.
Field days
Braving the cold rain and high winds of the North Atlantic summer, I found myself spending much of June, 2018, nose pressed against the coastal rocks, hand-lens at the ready, examining ancient trace fossils.
Accommodation on the island was limited for most of our time in Newfoundland, so most days we caught an early morning ferry from the mainland across the short stretch of the Labrador Sea to reach the island. The ferry trips gave us fantastic views of the island and its geology. The island has near-continuous exposure along the coastline, and even from a distance the sedimentary layering of the cliffs was visible, however many of the island’s outcrops are inaccessible without a boat.
We identified five main accessible field sites, some with fantastic coastal geography, including rock columns and bedding platforms extending out to sea. At each site we looked at the components of the fossil and trace fossil assemblages (communities of different species), as well as how these assemblages are distributed relative to one another, both in terms of their position in the stratigraphy and their relative locations on the bedding planes.
 Left, To a sedimentologist the island’s coastline holds a wealth of information, with the interlayered sands and muds preserving a record of the ancient tides. It also preserves a history of the island’s mining activity with bands of red ironstone, visible at the base of the stack, and rusted equipment and mine outlets dotted along the cliffs.
Left, To a sedimentologist the island’s coastline holds a wealth of information, with the interlayered sands and muds preserving a record of the ancient tides. It also preserves a history of the island’s mining activity with bands of red ironstone, visible at the base of the stack, and rusted equipment and mine outlets dotted along the cliffs.
Fossil assemblages
Broadly speaking, Bell Island’s fossils can be divided into three main assemblages (communities): microbial mats, simple shallow-burrowers and complex deep-burrowers. These assemblages may occur alone in a bedding plane or together, particularly in the case of the microbial mats and the shallow-burrowers.
The matgrounds are preserved either as wrinkle structures or as elephant skin fabrics. The two textures can be seen on the same bedding plane and can blend laterally into one another, indicating that they form part of the same matground community. These matgrounds may be crisscrossed by simple surface trails (trace fossils) that rarely penetrate deeper than a few mm into the sediment.
 Right, The microbial textures, wrinkle structures & elephant skin fabrics were often best preserved on the boundary between the sandstones and mudstones, with the microbial texture preserved on the top of the sands. As a result, outcrops like these containing multiple sandstone layers were some of the best places to find microbial textures.
Right, The microbial textures, wrinkle structures & elephant skin fabrics were often best preserved on the boundary between the sandstones and mudstones, with the microbial texture preserved on the top of the sands. As a result, outcrops like these containing multiple sandstone layers were some of the best places to find microbial textures.
The simple, shallow-burrowing assemblages are mostly made up of horizontally oriented burrows, which reach a maximum depth of 20 mm. More complex burrows are also present in these assemblages, albeit much rarer. These include U-shaped burrows, and larger, horizontally oriented, branching burrow networks. While the shallow-tier community can occur on its own on a bedding plane, it is more commonly found concentrated around the edge of, or directly beneath, the microbial mats. When in proximity to a matground, these shallow-tier communities form intense bioturbation zones—areas with a high burrow density.
Deeper-tier trace fossils are usually limited to layers well above the sandstones that preserve microbial textures (wrinkle structures and elephant skin fabrics). These deeper-tier communities can include simple, horizontally oriented burrows similar to those seen in the shallow-burrowing assemblage, however they also include vertically oriented burrows produced by organisms capable of deep burrowing and producing more complex burrow morphologies. These vertical burrows include simple non-branching forms and complex branching burrows produced by methodological sediment removal.
 Left, Wrinkle structures, here cross cut by simple fossils trails, (left) and Elephant skin fabrics (right) are microbial-induced sedimentary features produced by the binding of sediment (due to the secretion of a biological glue) and decay of a microbial matground
Left, Wrinkle structures, here cross cut by simple fossils trails, (left) and Elephant skin fabrics (right) are microbial-induced sedimentary features produced by the binding of sediment (due to the secretion of a biological glue) and decay of a microbial matground
Essential surface connections
Judging by these initial data, it seems that more complex, deep-tier burrowers were unable to survive in, or were less effective at colonising, matground sediments. So, the Precambrian matgrounds were likely to have initially been invaded by simpler, shallow-tier burrowers. Alternatively, the simple trace fossils we see associated with the matground may have been produced by deeper-tier burrowers that modified their burrowing behaviour in response to the presence of the matground. Either way, the trace fossils indicate that burrowing organisms that remain close to the sediment surface and the oxygenated water column are more likely to survive in, and effectively exploit, matground communities.
Close proximity to the sediment surface may have been the factor that allowed shallow burrowers to survive in the anoxic conditions and break-apart the matgrounds. By maintaining easy access to the oxygenated water column above, these animals could ‘dive’ into the anoxic sediment for brief periods of time, allowing them to feed before resurfacing for oxygen. They may also have been exploiting ‘waste’ oxygen produced by photosynthesising microorganisms in the matground itself (Meyer et al., Palaeo3 2014). By exploiting near surface oxygen, simpler organisms could have acted as one of the key pioneers in the Cambrian Substrate Revolution, breaking up the microbial sediment and opening up new ecological niches for more complex animals.
Today shallow-tier burrowers may be critical for preventing microbial matgrounds from re-establishing themselves on the modern seafloor as modern climate change continues to affect the global marine environment.
Above, Even in the overcast and rainy weather typical of the North Atlantic, the near continuous exposure of rock along Bell Island’s coastline is apparent from the mainland, with the sandstone-mudstone layering particularly visible on the right of the photograph.
Acknowledgments
Many thanks to my supervisors, Dr Liam Herringshaw, Dr Krysia Mazik and Prof Dan Parsons, at the University of Hull, and Dr Duncan McIlroy at the Memorial University of Newfoundland, and to the Geological Society of London for their support via the Elspeth Matthews Fund.
Affiliation
Catherine Mascord is a PhD Student at the University of Hull; e-mail:
[email protected]. Catherine was a 2018 recipient of the Geological Society of London’s Elspeth Matthews Fund.
Further reading
Buatois, L., et al. (2013) Global deglaciation and the re‐appearance of microbial matground‐dominated ecosystems in the late Paleozoic of Gondwana. Geobiology, 11(4): p. 307-317.
Crimes, T. and Droser, M. (1992) Trace Fossils and Bioturbation: The Other Fossil Record. The Annual Review of Ecology, Evolution, and Systematics, 23: p. 339-360.
Droser, M., Jensen, S. and Gehling, J. (2002) Trace Fossils and Substrates of the Terminal Proterozoic-Cambrian Transition: Implications for the Roecord of Early Bilaterians and Sediment Mixing. PNAS 99(20): p. 12572-12576.
Fedonkin, M. (1978) Ancient Trace Fossils and the Ways of Behavioral Evolution of Mud Eaters. Paleontological Journal, 12: p. 106-112.
Feng, X., et al. (2018) Unusual shallow marine matground-adapted benthic biofacies from the Lower Triassic of the northern Paleotethys: Implications for biotic recovery following the end-Permian mass extinction. Earth Science Reviews, In Press.
https://doi.org/10.1016/j.earscirev.2018.07.012
Gehling, J. (1999) Microbial Mats in Terminal Proterozoic Silicilastics: Ediacaran Death Masks. PALAIOS 14, 40-57.
Gerbersdorf, S., et al. (2009) Microbial Assemblages as Ecosystem Engineers. Journal of Soils and Sediments 9, 640-652.
Hotinski, R.M., et al., (2001) Ocean stagnation and end-Permian anoxia. Geology 29, 7-10.
Jensen, S. (2013) The Proterozoic and Earliest Cambrian Trace Fossil Record; Patterns, Problems and Perspectives. Integrative and Comparative Biology 43, 219-228.
King, A.F., (1998) Geology of the Avalon Peninsula, Newfoundland (parts of 1K, 1L, 1M, 1N and 2C). Newfoundland Department of Mines and Energy, Map 88 - 01.
Mata, S. and Bottjer, D. (2012) Microbes and Mass Extinctions: Paleoenvironmental Distribution of Microbialites During Times of Biotic Crisis. Geobiology. Special Issue: Microbes and Paleoenvironments 10, 3-24.
McIlroy, D. and Logan, G. (1999) The Impact of Bioturbation on Infaunal Ecology and Evolution During the Proterozoic-Cambrian Transition. PALAIOS 14, 58-72.
Meyer, M., et al., (2014) Interactions Between Edicaran Animals and Microbial Mats Insights from Lamonte trevallis, a New Trace Fossil from the Dengying formation of South China. Palaeogeography, Palaeoclimatology, Palaeoecology 396, 62-74.
Parsons, D., et al. (2016) The role of biophysical cohesion on subaqueous bed form size. Geophysical Research Letters 43, 1566-1573.
Pecoits, E., et al., (2012) Bilateran Burrows and Grazing Behaviour at >585 Million Years Ago. Science 336, p. 1693-1696.